Movie
Movie Controller
Controller

+ Open data
Open data

- Basic information
Basic information
| Entry | Database: PDB / ID: 7ook | ||||||||||||||||||
|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|
| Title | Bacteriophage PRD1 Major Capsid Protein P3 in complex with CPZ | ||||||||||||||||||
 Components Components | Major capsid protein P3 | ||||||||||||||||||
 Keywords Keywords |  VIRAL PROTEIN / protein complex / chlorpromazine (CPZ) VIRAL PROTEIN / protein complex / chlorpromazine (CPZ) | ||||||||||||||||||
| Function / homology | Bacteriophage PRD1, P3 / Bacteriophage PRD1, P3, N-terminal / P3 major capsid protein / Group II dsDNA virus coat/capsid protein / Viral coat protein subunit / viral capsid / Chem-Z80 / Major capsid protein P3 Function and homology information Function and homology information | ||||||||||||||||||
| Biological species |   Enterobacteria phage PRD1 (virus) Enterobacteria phage PRD1 (virus) | ||||||||||||||||||
| Method | X-RAY DIFFRACTION / SYNCHROTRON / MOLECULAR REPLACEMENT / Resolution: 2.23 Å | ||||||||||||||||||
 Authors Authors | Duyvesteyn, H.M.E. / Peccati, F. / Martinez-Castillo, A. / Jimenez-Oses, G. / Oksanen, H.M. / Stuart, D.I. / Abrescia, N.G.A. | ||||||||||||||||||
| Funding support |  Spain, Spain,  United Kingdom, 5items United Kingdom, 5items
| ||||||||||||||||||
 Citation Citation | Journal: Nanoscale / Year: 2021 Title: Bacteriophage PRD1 as a nanoscaffold for drug loading Authors: Duyvesteyn, H.M.E. / Santos-Perez, I. / Peccati, F. / Martinez-Castillo, A. / Walter, T.S. / Reguera, D. / Goni, F.M. / Jimenez-Oses, G. / Oksanen, H.M. / Stuart, D.I. / Abrescia, N.G.A. #1: Journal: Acta Crystallogr D Biol Crystallogr / Year: 2002Title: The X-ray crystal structure of P3, the major coat protein of the lipid-containing bacteriophage PRD1, at 1.65 A resolution. Authors: Stacy D Benson / Jaana K H Bamford / Dennis H Bamford / Roger M Burnett /  Abstract: P3 has been imaged with X-ray crystallography to reveal a trimeric molecule with strikingly similar characteristics to hexon, the major coat protein of adenovirus. The structure of native P3 has now ...P3 has been imaged with X-ray crystallography to reveal a trimeric molecule with strikingly similar characteristics to hexon, the major coat protein of adenovirus. The structure of native P3 has now been extended to 1.65 A resolution (R(work) = 19.0% and R(free) = 20.8%). The new high-resolution model shows that P3 forms crystals through hydrophobic patches solvated by 2-methyl-2,4-pentanediol molecules. It reveals details of how the molecule's high stability may be achieved through ordered solvent in addition to intra- and intersubunit interactions. Of particular importance is a 'puddle' at the top of the molecule containing a four-layer deep hydration shell that cross-links a complex structural feature formed by 'trimerization loops'. These loops also link subunits by extending over a neighbor to reach the third subunit in the trimer. As each subunit has two eight-stranded viral jelly rolls, the trimer has a pseudo-hexagonal shape to allow close packing in its 240 hexavalent capsid positions. Flexible regions in P3 facilitate these interactions within the capsid and with the underlying membrane. A selenometh-ionine P3 derivative, with which the structure was solved, has been refined to 2.2 A resolution (R(work) = 20.1% and R(free) = 22.8%). The derivatized molecule is essentially unchanged, although synchrotron radiation has the curious effect of causing it to rotate about its threefold axis. P3 is a second example of a trimeric 'double-barrel' protein that forms a stable building block with optimal shape for constructing a large icosahedral viral capsid. A major difference is that hexon has long variable loops that distinguish different adenovirus species. The short loops in P3 and the severe constraints of its various interactions explain why the PRD1 family has highly conserved coat proteins. #2: Journal: Cell / Year: 1999Title: Viral evolution revealed by bacteriophage PRD1 and human adenovirus coat protein structures. Authors: S D Benson / J K Bamford / D H Bamford / R M Burnett / Abstract: The unusual bacteriophage PRD1 features a membrane beneath its icosahedral protein coat. The crystal structure of the major coat protein, P3, at 1.85 A resolution reveals a molecule with three ...The unusual bacteriophage PRD1 features a membrane beneath its icosahedral protein coat. The crystal structure of the major coat protein, P3, at 1.85 A resolution reveals a molecule with three interlocking subunits, each with two eight-stranded viral jelly rolls normal to the viral capsid, and putative membrane-interacting regions. Surprisingly, the P3 molecule closely resembles hexon, the equivalent protein in human adenovirus. Both viruses also have similar overall architecture, with identical capsid lattices and attachment proteins at their vertices. Although these two dsDNA viruses infect hosts from very different kingdoms, their striking similarities, from major coat protein through capsid architecture, strongly suggest their evolutionary relationship. #3: Journal: Nature / Year: 2004Title: Insights into assembly from structural analysis of bacteriophage PRD1. Authors: Nicola G A Abrescia / Joseph J B Cockburn / Jonathan M Grimes / Geoffrey C Sutton / Jonathan M Diprose / Sarah J Butcher / Stephen D Fuller / Carmen San Martín / Roger M Burnett / David I ...Authors: Nicola G A Abrescia / Joseph J B Cockburn / Jonathan M Grimes / Geoffrey C Sutton / Jonathan M Diprose / Sarah J Butcher / Stephen D Fuller / Carmen San Martín / Roger M Burnett / David I Stuart / Dennis H Bamford / Jaana K H Bamford / Abstract: The structure of the membrane-containing bacteriophage PRD1 has been determined by X-ray crystallography at about 4 A resolution. Here we describe the structure and location of proteins P3, P16, P30 ...The structure of the membrane-containing bacteriophage PRD1 has been determined by X-ray crystallography at about 4 A resolution. Here we describe the structure and location of proteins P3, P16, P30 and P31. Different structural proteins seem to have specialist roles in controlling virus assembly. The linearly extended P30 appears to nucleate the formation of the icosahedral facets (composed of trimers of the major capsid protein, P3) and acts as a molecular tape-measure, defining the size of the virus and cementing the facets together. Pentamers of P31 form the vertex base, interlocking with subunits of P3 and interacting with the membrane protein P16. The architectural similarities with adenovirus and one of the largest known virus particles PBCV-1 support the notion that the mechanism of assembly of PRD1 is scaleable and applies across the major viral lineage formed by these viruses. #4: Journal: Nature / Year: 2004 Title: Membrane structure and interactions with protein and DNA in bacteriophage PRD1. Authors: Joseph J B Cockburn / Nicola G A Abrescia / Jonathan M Grimes / Geoffrey C Sutton / Jonathan M Diprose / James M Benevides / George J Thomas / Jaana K H Bamford / Dennis H Bamford / David I Stuart / Abstract: Membranes are essential for selectively controlling the passage of molecules in and out of cells and mediating the response of cells to their environment. Biological membranes and their associated ...Membranes are essential for selectively controlling the passage of molecules in and out of cells and mediating the response of cells to their environment. Biological membranes and their associated proteins present considerable difficulties for structural analysis. Although enveloped viruses have been imaged at about 9 A resolution by cryo-electron microscopy and image reconstruction, no detailed crystallographic structure of a membrane system has been described. The structure of the bacteriophage PRD1 particle, determined by X-ray crystallography at about 4 A resolution, allows the first detailed analysis of a membrane-containing virus. The architecture of the viral capsid and its implications for virus assembly are presented in the accompanying paper. Here we show that the electron density also reveals the icosahedral lipid bilayer, beneath the protein capsid, enveloping the viral DNA. The viral membrane contains about 26,000 lipid molecules asymmetrically distributed between the membrane leaflets. The inner leaflet is composed predominantly of zwitterionic phosphatidylethanolamine molecules, facilitating a very close interaction with the viral DNA, which we estimate to be packaged to a pressure of about 45 atm, factors that are likely to be important during membrane-mediated DNA translocation into the host cell. In contrast, the outer leaflet is enriched in phosphatidylglycerol and cardiolipin, which show a marked lateral segregation within the icosahedral asymmetric unit. In addition, the lipid headgroups show a surprising degree of order. | ||||||||||||||||||
| History |
|
- Structure visualization
Structure visualization
| Structure viewer | Molecule: MolmilJmol/JSmol |
|---|
- Downloads & links
Downloads & links
-Download
| PDBx/mmCIF format | 7ook.cif.gz | 519.9 KB | Display | PDBx/mmCIF format |
|---|---|---|---|---|
| PDB format | pdb7ook.ent.gz | 352.8 KB | Display | PDB format |
| PDBx/mmJSON format | 7ook.json.gz | Tree view | PDBx/mmJSON format | |
| Others |  Other downloads Other downloads |
-Validation report
| Arichive directory | https://data.pdbj.org/pub/pdb/validation_reports/oo/7ookftp://data.pdbj.org/pub/pdb/validation_reports/oo/7ook | HTTPS FTP |
|---|
-Related structure data
| Related structure data |  1hx6S S: Starting model for refinement C: citing same article ( |
|---|---|
| Similar structure data |

-Links
 PDBj
PDBj

- Assembly
Assembly
| Deposited unit | 
| ||||||||||||
|---|---|---|---|---|---|---|---|---|---|---|---|---|---|
| 1 |
| ||||||||||||
| Unit cell |
|
-Components
-Protein , 1 types, 3 molecules CBA
| #1: Protein | Mass: 43477.414 Da / Num. of mol.: 3 / Source method: isolated from a natural source / Source: (natural) Enterobacteria phage PRD1 (virus) / References: UniProt: P22535 |
|---|
-Non-polymers , 5 types, 435 molecules 



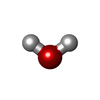
| #2: Chemical | ChemComp-NA /  Mass: 22.990 Da / Num. of mol.: 5 / Source method: isolated from a natural source / Formula: Na Mass: 22.990 Da / Num. of mol.: 5 / Source method: isolated from a natural source / Formula: Na#3: Chemical | ChemComp-MPD / ( 2-Methyl-2,4-pentanediol Mass: 118.174 Da / Num. of mol.: 10 / Source method: obtained synthetically / Formula: C6H14O2 / Comment: precipitant*YM Mass: 118.174 Da / Num. of mol.: 10 / Source method: obtained synthetically / Formula: C6H14O2 / Comment: precipitant*YM#4: Chemical | Chlorpromazine Mass: 318.864 Da / Num. of mol.: 3 / Source method: isolated from a natural source / Formula: C17H19ClN2S / Feature type: SUBJECT OF INVESTIGATION / Comment: medication, antipsychotic*YM Mass: 318.864 Da / Num. of mol.: 3 / Source method: isolated from a natural source / Formula: C17H19ClN2S / Feature type: SUBJECT OF INVESTIGATION / Comment: medication, antipsychotic*YM#5: Chemical | Chloride Mass: 35.453 Da / Num. of mol.: 3 / Source method: obtained synthetically / Formula: Cl Mass: 35.453 Da / Num. of mol.: 3 / Source method: obtained synthetically / Formula: Cl#6: Water | ChemComp-HOH / | WaterMass: 18.015 Da / Num. of mol.: 414 / Source method: isolated from a natural source / Formula: H2O |
|---|
-Details
| Has ligand of interest | Y |
|---|
-Experimental details
-Experiment
| Experiment | Method: X-RAY DIFFRACTION / Number of used crystals: 1 |
|---|
- Sample preparation
Sample preparation
| Crystal | Density Matthews: 3.66 Å3/Da / Density % sol: 66.41 % |
|---|---|
| Crystal grow | Temperature: 293 K / Method: vapor diffusion, sitting drop / Details: please see: PMID: 10499799 |
-Data collection
| Diffraction | Mean temperature: 100 K / Serial crystal experiment: N |
|---|---|
| Diffraction source | Source: SYNCHROTRON / Site: Diamond / Beamline: I24 / Wavelength: 0.9686 Å |
| Detector | Type: DECTRIS PILATUS3 6M / Detector: PIXEL / Date: Jun 4, 2018 |
| Radiation | Protocol: SINGLE WAVELENGTH / Monochromatic (M) / Laue (L): M / Scattering type: x-ray |
| Radiation wavelength | Wavelength: 0.9686 Å / Relative weight: 1 |
| Reflection | Resolution: 2.02→54.5 Å / Num. obs: 120623 / % possible obs: 99.97 % / Redundancy: 13.1 % / Biso Wilson estimate: 42.64 Å2 / CC1/2: 0.999 / Net I/σ(I): 14.03 |
| Reflection shell | Resolution: 2.02→2.092 Å / Redundancy: 13.3 % / Num. unique obs: 11888 / CC1/2: 0.595 / CC star: 0.864 / % possible all: 100 |
- Processing
Processing
| Software |
| |||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||
|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|
| Refinement | Method to determine structure: MOLECULAR REPLACEMENT Starting model: 1hx6 Resolution: 2.23→39.83 Å / SU ML: 0.2694 / Cross valid method: FREE R-VALUE / σ(F): 1.37 / Phase error: 20.4506 Stereochemistry target values: GeoStd + Monomer Library + CDL v1.2
| |||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||
| Solvent computation | Shrinkage radii: 0.9 Å / VDW probe radii: 1.11 Å / Solvent model: FLAT BULK SOLVENT MODEL | |||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||
| Displacement parameters | Biso mean: 49.23 Å2 | |||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||
| Refinement step | Cycle: LAST / Resolution: 2.23→39.83 Å
| |||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||
| Refine LS restraints |
| |||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||
| LS refinement shell |
|