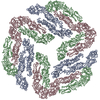
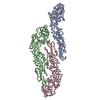
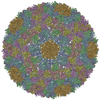
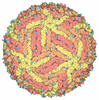
ジャーナル: J Virol / 年: 2013 タイトル: Structural changes in dengue virus when exposed to a temperature of 37°C. 著者: Guntur Fibriansah / Thiam-Seng Ng / Victor A Kostyuchenko / Jaime Lee / Sumarlin Lee / Jiaqi Wang / Shee-Mei Lok / 要旨: Previous binding studies of antibodies that recognized a partially or fully hidden epitope suggest that insect cell-derived dengue virus undergoes structural changes at an elevated temperature. This ...Previous binding studies of antibodies that recognized a partially or fully hidden epitope suggest that insect cell-derived dengue virus undergoes structural changes at an elevated temperature. This was confirmed by our cryo-electron microscopy images of dengue virus incubated at 37°C, where viruses change their surface from smooth to rough. Here we present the cryo-electron microscopy structures of dengue virus at 37°C. Image analysis showed four classes of particles. The three-dimensional (3D) map of one of these classes, representing half of the imaged virus population, shows that the E protein shell has expanded and there is a hole at the 3-fold vertices. Fitting E protein structures into the map suggests that all of the interdimeric and some intradimeric E protein interactions are weakened. The accessibility of some previously found cryptic epitopes on this class of particles is discussed.
 ムービー
ムービー コントローラー
コントローラー

 データを開く
データを開く
 基本情報
基本情報 要素
要素 キーワード
キーワード VIRUS (ウイルス) / BREATHING STRUCTURE / DYNAMIC STRUCTURE
VIRUS (ウイルス) / BREATHING STRUCTURE / DYNAMIC STRUCTURE 機能・相同性情報
機能・相同性情報
 データ登録者
データ登録者 引用
引用
 構造の表示
構造の表示 ダウンロードとリンク
ダウンロードとリンク その他のダウンロード
その他のダウンロード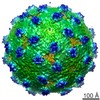







 PDBj
PDBj
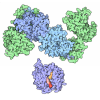
 集合体
集合体